|
Cambios
en la composición y estructura de la vegetación
arbórea en un área impactada por la ganadería en
el corregimiento de Santa Cecilia,
Pueblo Rico, Risaralda Changes in the composition and structure of the arboreal vegetation in an area impacted by the livestock in the corregimiento of Santa Cecilia, Pueblo Rico, Risaralda Luis Javier Mosquera Ramos1, Reimer Rengifo Ibargüen1, Omar Quinto Mosquera2 1 Instituto de Investigaciones Ambientales del Pacífico “John Von Neumann”, grupo de investigación Conocimiento, manejo y conservación de los ecosistemas del Chocó Biogeográfico. e-mail: lmosquera@iiap.org.co e-mail: rrengifo@iiap.org.co 2 Instituto de Investigaciones Ambientales del Pacífico “John Von Neumann”. Autor correspondencia: e-mail: omar.quint13@hotmail.com Recepción: Septiembre 5, 2017 Aprobación: Noviembre 29, 2017 Editor asociado: Ramírez G. DOI: https://doi.org/10.51641/bioetnia.v15i1.195 Resumen Con fin de determinar los cambios en la composición y estructura de la vegetación arbórea en un área impactada (75 ha) por ganadería durante el período 2015-2017, se muestreó y monitoreó todos los individuos de porte arbóreo presente en una parcela de 0,25 ha (50x50 m). Los muestreos realizados, permitieron registrar 199 individuos pertenecientes a 14 familias y 19 especies. Las familias mejor representadas en cuanto a riqueza de especies, fueron: Moraceae con 3, seguida de Malvaceae, Melastomataceae y Urticaceae con 2 especies cada una. A nivel de especies, sobresalen por su abundancia Vismia angustifolia (punta de lanza) con 12 individuos, Jacaranda sp. (gualanday o pinguasí) y Lacistema aggregatum (café de monte) con 9 individuos cada una, e Himatanthus articulatus (mamancallo) con 8 individuos. La composición de especies fue similar a lo largo de los muestreos, sin embargo, en el año 2016 se registró la especie Apeiba membranacea y Brosimum utile en el año 2017. El promedio diamétrico de los individuos osciló entre 18 y 19 cm. La dinámica de crecimiento de los árboles muestra, que a pesar de que el bosque no ha alcanzado su madurez, los individuos de las clases más altas, disminuyen su tasa de crecimiento al paso de los años. De otra parte, la mortalidad y reclutamiento varió entre los períodos evaluados. Para el período 2015-2016, no se presentó mortalidad, sin embargo, se reclutaron 10 individuos, para un total de 65 árboles para este período, representando un incremento de 0,363 m2, equivalente al 16% del área basal con respecto al año 2015 (1.897 m2). A pesar de la poca variación en los parámetros estructurales y de composición, las familias y especies aumentan, conforme avanza el estadío sucesional, demostrando la tendencia de recuperación de atributos de diversidad, composición y funcionalidad del ecosistema. Palabras clave: Composición y estructura, Ganadería, Santa Cecilia, Vegetación. Abstract In order to determine the changes in the composition and structure of the arboreal vegetation in an impacted area (75 ha) by livestock during the period 2015-2017, all the individuals of arboreal size present in a plot of 0.25ha (50x50 m). The samplings carried out allowed to register 199 individuals belonging to 14 families and 19 species. The best represented families in terms of species richness were: Moraceae with 3, followed by Malvaceae, Melastomataceae and Urticaceae with 2 species each. At the species level, Vismia angustifolia (Spearhead) with 12 individuals, Jacaranda sp. (gualanday or pinguasí) and Lacistema aggregatum (mountain coffee) with 9 individuals each, and Himatanthus articulatus (mamancallo) with 8 individuals. The species composition was similar throughout the samplings, however, in 2016 the species Apeiba membranacea was recorded, and Brosimum utile in 2017. The diametric average of the individuals ranged between 18 and 19 cm. The growth dynamics of the trees shows that despite the fact that the forest has not reached its maturity, the individuals of the highest classes decrease their growth rate over the years. On the other hand, mortality and recruitment varied between the evaluated periods. For the period 2015-2016, there was no mortality, however, 10 individuals were recruited, for a total of 65 trees for this period, representing an increase of 0.363 m2, equivalent to 16% of the basal area with respect to 2015 (1.897 m2). Despite the little variation in the structural and composition parameters, the families and species obtained as the successional stage progresses, demonstrating the tendency of recovery of attributes of diversity, composition and functionality of the ecosystem. Keywords: Composition and structure, Livestock, Santa Cecilia, Vegetation. Introducción Los ecosistemas tropicales son los más biodiversos del planeta (Calle 1994); sin embargo, en las últimas décadas, estos se han vistos afectados por diversas actividades antrópicas, donde la ganadería resulta ser una de estas más impactantes (Geist y Lambin 2001, 2002), pues implica la tala de árboles, ocasiona compactación de los suelos, deterioro y pérdida en la capacidad productiva del mismo, afecta la dinámica del ecosistema, así como los bienes y servicios que estos prestan a las comunidades humanas (González et al. 2011). En Colombia, esta realidad se extiende por todo el territorio y en muchos casos han dado paso a la formación de bosques secundarios, que en la mayoría de las veces son el resultado de tierras abandonadas después de su uso prolongado (Carim et al. 2007). La transformación del bosque en este territorio inicia con la remoción de pequeñas áreas que se usan para la agricultura de subsistencia. Después, estas áreas son reemplazadas por pastizales para ganadería. Con el paso de los años, algunas de estas áreas son abandonadas debido a la baja productividad del suelo, migración rural-urbana, mejoras en la tecnología y la globalización de mercados (Sánchez-Cuervo et al. 2012). Caso como el descrito antes, se presenta en el corregimiento de Santa Cecilia, Risaralda, donde, producto de la ganadería, existen grandes extensiones de tierras con baja diversidad vegetal y sobrepastoreo del ganado, que afectan las funciones ecosistémicas que dependen directamente de la presencia del componente arbóreo tales como: la captura o secuestro de CO2, regulación hídrica, del clima y control de la erosión entre otras. Por otra parte, y teniendo en cuenta que la recuperación del ecosistema depende tanto del tipo de disturbio como la intensidad de la afectación, es necesario desarrollar procesos investigativos que permitan comprender la dinámica y funcionamiento del bosque de manera que se puedan dimensionar los cambios a través del tiempo. Desde esta perspectiva vale la pena resaltar las investigaciones realizadas por Janzen (1970), Gentry (1982), Clark y Clark (1984), Connell (1978), Brown y Lugo (1990), y Lieberman et al. (1995), quienes se han esforzado por conocer la dinámica de estas áreas disturbadas, tratando de dilucidar los complejos procesos que gobiernan la dinámica, la regeneración y la consecuente permanencia de las especies. También vale la pena resaltar los esfuerzos investigativos realizados por Bonilla et al. (2011), Ordoñez (2016), IIAP (2017), Valois y Martínez (2017), los cuales se enfocaron en comprender la dinámica de recuperación de bosques impactados por minería y la tala selectiva de árboles en el andén Pacífico colombiano. Lo anterior evidencia el vacío de información existente en torno a los ecosistemas impactados por la ganadería. De allí que con esta investigación se pretende evaluar los cambios ocurridos en la composición y estructura de la vegetación arbórea en un área impactada por la ganadería en el corregimiento de Santa Cecilia, entre los años 2015-2017, con el fin de generar información confiable que ayude a comprender la dinámica sucesional del ecosistema luego de este tipo de disturbio. Metodología Área de estudio. Se realizó en el corregimiento de Santa Cecilia, ubicado al noroccidente del departamento de Risaralda, a 34 km de la cabecera municipal de Pueblo Rico, sobre la vía que conduce al departamento del Chocó. Esta zona se caracteriza por presentar un clima cálido per húmedo, con temperatura promedio anual mayor de 24°C, altura entre 370 y 1000 msnm, un índice de humedad mayor a 100, precipitaciones entre los 2.000 a 4.000 mm al año, vegetación compuesta por pastos y cultivos en rastrojados, contiene algunos elementos de la vegetación natural, por el sistema de cultivo desarrollado tradicionalmente (Duque 1997). La parcela de restauración pasiva está ubicada en jurisdicción del área de majeño especial Amurrupá, sobre la margen derecha del río Amurrupá, en el sitio conocido localmente como Las Colonias, en inmediaciones del sendero El Volga que conduce al Alto Amurrupá. Geográficamente se localiza a los 05°20´12,´´ N y 076°09´19,´´W, a una altura de 420 msnm. Este sitio, por más de 30 años, fue destinado a la actividad ganadera, sin embargo, para la fecha del primer muestreo tenía aproximadamente 16 años de haber sido abandonado y en la actualidad se encuentra en proceso de recuperación natural (Figura 1). Figura 1. Localización del corregimiento de Santa Cecilia, Risaralda
 Fuente: CARDER (2012) Método
Toma de datos. Para la caracterización y posterior monitoreo de la vegetación de porte arbóreo, en el año 2015 se estableció una parcela cuadrada de 50x50 m (0,25 ha), la que a su vez fue dividida en 25 subparcelas de 10x10 m (100 m2) cada una, las cuales fueron enumeradas e identificadas a través del par ordenado (A1; A2, A3; B1, B2, B3; C1, C2… etc.). Al interior de ellas, se censaron todos los individuos con DAP≥10 cm, a los que, además se le estimó la altura a la primera rama (comercial), altura total, y se tomó información sobre usos y formas de vida o bioformas árbol, arbustos, helecho arbóreo y palmas entre otros (Villarreal et al. 2004). Para el monitoreo además de lo realizado en el año 2015, se determinó la mortalidad y reclutamiento para todos los individuos existentes, siguiendo el protocolo de Phillips et al. (2009). Se incluyeron como renuevos o reclutas todos los árboles que entraron a la categoría de tamaño mayor o igual a 10 cm de DAP. Se colectaron entre dos y tres ejemplares de cada especie o morfo especie, en lo posible en estado fértil. Este material fue preparado de acuerdo con las técnicas o métodos estandarizados. La identificación taxonómica se realizó por comparaciones con catálogos, monografías y literaturas especializadas o guías de flora, tales como: Gentry (1993), Quiñones (2001), Salinas y Betancur (2005), Mendoza et al. (2004), Cárdenas (2007), Cárdenas et al. (2008), Cardona et al. (2010), Cardona et al. (2011), Peñuela y Jiménez (2010), Alzate et al. (2012), Díaz y Elcoro (2009). Análisis de los datos. El análisis de la estructura horizontal del ecosistema, se realizó mediante el cálculo de la densidad total, frecuencia relativa, áreas basales, área basal total e IVI (Índice de Valor de Importancia); todas estas variables estructurales se calcularon siguiendo los lineamientos recomendados por Otavo (2002), mientras que para la caracterización vertical se siguió la propuesta de Rangel y Lozano (1986), citada en Villarreal et al. (2004), que propone los siguientes estratos, según su altura total: rasante (<0,3 m), herbáceo (0,3-1,5 m), arbustivo (1,5-5 m), subarbóreo o de arbolitos (5-12 m), arbóreo inferior (12-25 m) y arbóreo superior (>25m). Análisis de los cambios en los parámetros estructurales entre muestreos. Para la medición de parámetros demográficos como mortalidad (m) y reclutamiento (r), se utilizaron las siguientes fórmulas:  donde: N15 = número de individuos presentes en el 2015 NS = número de individuos sobrevivientes en el 2017 NS = N15- = número de individuos muertos en el período) N15 = NS+ = número de individuos reclutados en el tiempo (T) BA15 = área basal de los individuos presentes en el 2015 Bas15 = área basal de los individuos en el año 2015 y presentes en el 2017 BAs17 = basal en el 2017 de los individuos presentes en el 2017 BA17 = BAs17 + = el área basal de los individuos reclutados en el 2017 Para el análisis de crecimiento se calcularon los incrementos medios anuales (IMA) únicamente con las mediciones de los individuos presentes entre el intervalo de los años 2015 y 2017, porque las diferencias de mediciones de año a año son tan pequeñas que es difícil obtener valores precisos (Murphy y Lugo 1986). Resultados y discusiones Composición florística. Durante los muestreos realizados entre los años 2015 y 2017, se registraron 199 individuos (55 en el 2015, 65 en 2016 y 76 en 2017) pertenecientes a 14 familias y 19 especies (Anexo a). Entre las familias mejor representadas en cuanto a riqueza, se destaca Moraceae con 3 especies, seguida de Malvaceae, Melastomataceae y Urticaceae con 2 especies cada una. Estas familias se mantuvieron como las más representativas en los tres muestreos. Las familias botánicas aquí registradas son un poco similares a las reportadas en otras investigaciones realizadas en bosques secundarios afectados por distintas actividades productivas. Bonilla et al. (2011), reportan a Melastomataceae, Clusiaceae, Apocynaceae, Myristicaceae y Lecythidaceae como las familias dominantes en bosque afectado por el aprovechamiento forestal. Por su parte, Moraes et al. (2002), reportan a Fabaceae, Rubiaceae, Moraceae, Meliaceae, y Malvaceae como las familias más diversas en los bosques secundarios del municipio de San Carlos, Nicaragua. En general, las familias identificadas en este estudio se pueden considerar típicas de bosques tropicales secundarios, pues diversas investigaciones corroboran su ocurrencia en este tipo de ecosistemas (Finegan 1992, 1996, Sevegnani y Baptista 1996, Finegan y Delgado 2000, Steininger 2000). A nivel de especies, sobresalen por su abundancia Vismia angustifolia (punta de lanza) con 12 individuos, Jacaranda sp. (gualanday o pinguasí) Lacistema aggregatum (café de monte) con 9 individuos cada una, e Himatanthus articulatus (mamancallo) con 8 individuos. La composición de especies fue bastante similar a lo largo de los muestreos, sin embargo, en el año 2016 se registró la especie Apeiba membranacea, mientras que para el año 2017, se incorporó Brosimum utile.  En cuanto al número de individuos por especies, se observó poca variación (entre 1 y 3), pues las especies que se reportaron como las dominantes en el año 2015, fueron constantes hasta el muestreo del 2017. Los mayores aumentos se presentaron en las especies Jacaranda sp., Vismia angustifolia, Miconia sp., Himatanthus articulatus y Socratea exorrhiza (Anexo a). Lo anterior demuestra la capacidad que tienen estas especies para sobrevivir en estos ambientes disturbados por la práctica ganadera. Por otra parte, en el período 2015 y 2016, cinco especies (Tessaria integrifolia, Himatanthus articulatus, Socratea exorrhiza, Jacaranda sp., Lacistema aggregatum) mostraron incrementos de 1,5 individuos en promedio. De igual manera, se destaca la incorporación de la especie Apeiba membranacea con un individuo, el cual persistió hasta el muestreo del año 2017. En cuanto al ciclo 2016-2017, se resalta a Socratea exorrhiza, Jacaranda sp., Vismia angustifolia, Ocotea sp., Bellucia axinanthera, Miconia sp., Ficus insipida y Cecropia angustifolia como las especies que aumentaron sus poblaciones, con un promedio similar al registrado en el período inmediatamente anterior. Socratea exorrhiza, Jacaranda sp. y Miconia sp. fueron las únicas especies que mostraron un constate aumento en el número de individuos a lo largo de los muestreos, lo que denota su dinámica de crecimiento y adaptación a este ambiente (Figura 2). Figura 2. Especies de vegetales de porte arbóreo más abundante período 2015-2017 en la parcela
de restauración pasiva ubicada en el corregimiento de Santa Cecilia (Risaralda)  Estructura diamétrica. El promedio diamétrico de los individuos varió a lo largo de los muestreos realizados, pasando de 18,3 cm en 2015 a 18,5 cm en 2016 y 19 cm en el 2017. En el año 2015, los mayores promedios diamétrico los presentaron las especies Vismia angustifolia (58,9 cm), Ocotea sp. (33,7 cm), Tessaria integrifolia (33,1 cm) e Inga sp. (33,1 cm). Para el año 2016 sobresalió Cecropia angustifolia (69,1 cm), Bellucia axinanthera (48,1 cm), Miconia sp. (44,9 cm), Cecropia sp. (136 cm) y Ocotea sp. (43,3 cm); mientras que para el año 2017 lo fueron Cecropia angustifolia (53,2 cm), Ocotea sp. (42,6 cm), Vismia angustifolia (40,1 cm), Miconia sp. (38,8 cm), y Bellucia axinanthera (36,3 cm). Para el caso de las especies V. angustifolia, C. angustifolia, B. axinanthera y Miconia sp., se observó un comportamiento atípico reflejado en la reducción del promedio diamétrico, lo que se puede atribuir a la desaparición o muerte de algunos individuos. Por otra parte, es necesario precisar que, de estas especies, Ocotea sp. sobre salió como la de mayor aporte diamétrico durante los tres muestreos, lo que pudo obedecer a la incorporación de un nuevo individuo, con 11 cm de diámetro. De otro lado, durante los tres muestreos realizados, la agrupación de los de individuos por clase diamétrica, mostró alta concentración de individuos en las clases menores con reducción acentuada para las clases mayores, representando la distribución en forma de J invertida (Figura 3), es decir, que hay un menor número de individuos en las clases superiores (DAP>20 cm). Esta distribución representa la tendencia del bosque en buscar la fase de homeostasis (equilibrio entre lo que se muere y lo que crece), aspectos representados por los movimientos que se presentan en el paso de individuos entre las clases diamétricas, lo cual es parte de la dinámica natural del bosque, en donde la cantidad de individuos que logran establecerse durante los primeros años va disminuyendo conforme aumenta la clase diamétrica producto de la competencia intra e interespecífica y de las exigencias lumínicas que requieren algunas de las especies para obtener un sitio dentro del bosque (Rodríguez y Brenes 2009). Al respecto, Orozco y Brumér (2002), manifiestan que este tipo de comportamiento refleja la presencia de un bosque secundario discetáneo o irregular. Sin embargo, para algunos autores (Arruda et al. 2011, Hernández-Stefanoni et al. 2011), la tendencia de la curva en forma de J invertida también señala que la comunidad vegetal se encuentra en coherente proceso de desarrollo en dirección a etapas de crecimiento y productividad vegetal más avanzados, una vez que se confirma la existencia de abundantes individuos jóvenes que irán a suceder individuos arbóreos pertenecientes a las clases diamétricas superiores. Figura 3. Distribución diamétrica de las especies arbóreas registradas en la parcela de
restauración pasiva de Santa Cecilia, Risaralda, durante el período 2015-2017 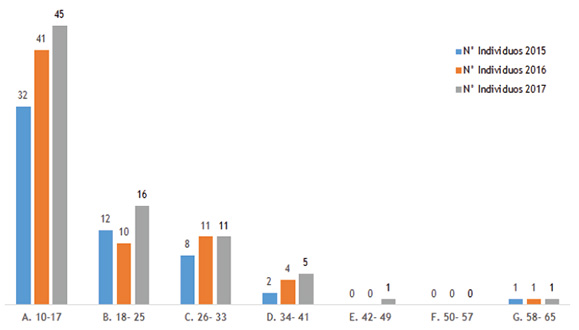 Para Newton (2007), este tipo de estructura dominada por muchos árboles pequeños sugiere que el rodal ha sufrido una reciente regeneración o establecimiento, después de un evento de disturbio. Al respecto Lamprecht (1990), manifiesta que la estructura disetánea, característica de este tipo de distribuciones diamétricas, es la mejor garantía de la supervivencia de la comunidad forestal, ya que los individuos de tallas superiores son eliminados ocasionalmente y sustituidos sin dificultad por individuos de las categorías diamétricas inferiores. De otro lado, al analizar el comportamiento de los individuos al interior de las clases diamétricas a lo largo de los muestreos (Figura 3), se aprecia que la clase A (10-17 cm), mostró un patrón de aumento logarítmico del número de individuos. Mientras en la clase B, se nota un cambio de patrón debido a la muerte de individuos en el año 2016. Para el caso de las clases superiores (F y G), no se presentó muerte ni reclutamiento, lo que significa que la abundancia fue constante. Estructura vertical. La vegetación arbórea registrada en la parcela de restauración pasiva, presentó una altura promedio de 13,4 m/ind, sobresaliendo Ocotea sp., Inga sp. y Castilla elastica como las especies más altas con 28, 24 y 24 m respectivamente; mientras que Brosimum utile, Vismia angustifolia e Hymatanthus articulata se registraron como las de menos altura con 6, 6,4 y 7 m en su orden. La Figura 4 muestra la distribución de árboles por clases de altura, donde se aprecia un patrón de crecimiento en las primeras cuatro clases, donde la B (10-12 m de altura) congrega la mayor cantidad de individuos pasando de 16 árboles en 2015 a 24 en 2016 y 33 en 2017, mostrando así, un aumento de más del 50% de individuos en relación con el año anterior en esta clase; sin embargo, a medida que aumenta el rango de altura y el período de muestreo, se percibe una disminución de individuos en dichas clases hasta del 28% (clase E). La dinámica de crecimiento de los árboles en los tres períodos, muestra que a pesar de que el bosque no ha alcanzado su madurez, los árboles de las clases más altas, disminuyen su tasa de crecimiento al paso de los años. Figura 4. Distribución por clase de altura de los individuos registrados en la parcela permanente
de restauración pasiva en Santa Cecilia, Risaralda, durante el período 2015-2017 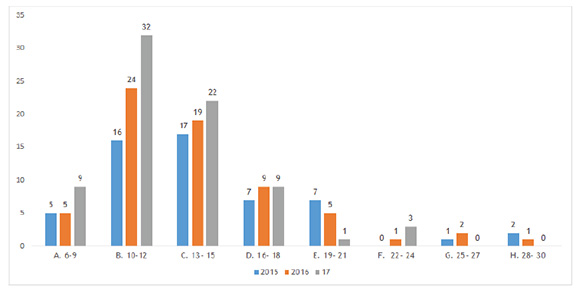 Por otra parte, de acuerdo con el arreglo de clases de alturas propuesto por Rangel y Velázquez (1997), los árboles aquí registrados, se distribuyen en tres estratos, donde la mayor cantidad de individuos (41: 54%) se ubican en el estrato subarbóreo o de arbolitos (5-12 m), siendo Hymatanthus articulata y Vismia angustifolia las especies más dominantes. También, 44% (34 individuos) se ubican en el estrato arbóreo inferior (12-25 m) con representación de las especies Vismia angustifolia, Jacaranda sp., Tessaria integrifolia, Miconia sp., Socratea exorrhiza y Cecropia sp., mientras en el estrato arbóreo superior (>25 m) se reporta un solo individuo de la especie Ocotea sp., pues de los tres individuos existentes tanto en el año 2015 como en el 2016, dos se registraron en este monitoreo como muertos, resultando así, solo la especie Ocotea sp. en el estrato emergente del bosque. Vismia angustifolia mostró la tasa más altas de crecimiento en el último año, seguida de Himatanthus articulatus con un promedio cerca de 2 cm de altura/año; estas especies, junto con Lacistema aggregatum, además de presentar la mayor cantidad de individuos en las diferentes clases de altura, también estuvieron presente en el mayor número de estas clases (5); mientras Jacaranda sp., Ocotea sp., y Tessaria integrifolia a diferencia de las anteriores, ocupan menor número de clases de altura a pesar de estar entre las más abundantes. Índice de valor de importancia de las especies (IVI). La Figura 5 muestra el comportamiento del peso o importancia ecológica de las especies durante el período 2015-2017. En ella se puede apreciar que para el año 2015 las especies ecológicamente más importante fueron: Vismia angustifolia 46,9 (15,6%), Ocotea sp. 30,2 (9,6%), Himatanthus articulatus 28,9 (10,1%), Lacistema aggregatum 28,1 (9,3%), Inga sp. 27 (9%). En los muestreos posteriores, estas especies continúan siendo las de mayor peso ecológico, junto con Jacaranda sp. 27 (9%). Esto a pesar de las variaciones presentadas en atributos como el área basal y la abundancia. Figura 5. Comportamiento de índice de valor de importancia ecológica de las especies
arbóreas registradas en la parcela de restauración pasiva de Santa Cecilia, Risaralda, durante el período 2015-2017  Para el caso de Vismia angustifolia, Himatanthus articulatus, Lacistema aggregatum el IVI, estuvo influido por la abundancia y la frecuencia. A diferencia de lo ocurrido con Ocotea sp., cuya importancia ecológica estuvo determinada por su área basal y la permanente ocurrencia en las unidades de muestreo (Anexos b, c y d). 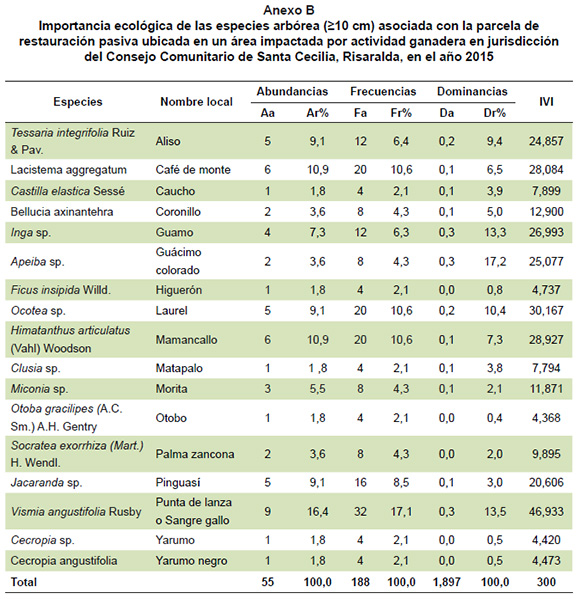   Mortalidad y reclutamiento. Varió entre los períodos evaluados. Para el ciclo 2015-2016, no se presentó mortalidad, sin embargo, se reclutaron 10 individuos, para un total de 65 árboles para este período, representando un incremento de 0,363 m2, equivalente al 16% del área basal con respecto al año 2015 (1.897 m2). Es necesario precisar que los individuos reclutados pertenecen a las especies Tessaria integrifolia, Himatanthus articulatus, Socratea exorrhiza, Jacaranda sp., Lacistema aggregatum y Miconia sp., donde la mayor cantidad de individuos los aportó Lacistema aggregatum (café de monte), lo que influyó en su abundancia para el muestreo de 2016. Con respecto al período 2016-2017, la mortalidad fue de 6 árboles (9,2%) correspondientes a las especies V. angustifolia, C. angustifolia, B axinanthera y Miconia sp., donde el patrón de mortalidad principal fue: muerte en pie, 3 árboles (50%); volcamiento, 2 (33,3%); y aprovechado 1 individuo (16,6%). Este patrón difiere con lo reportado por Quinto et al. (2009), quienes reportan el volcamiento de raíz como la causa de mortalidad más frecuente (25%). Por otra parte, el reclutamiento correspondiente a este mismo período, fue del 25% (20 árboles), representando un área basal de 0,719 m2, equivalente a 24,3% del área basal registrada en el 2017 (2.961m2). De los individuos reclutados, la mayor cantidad fue para las especies Jacaranda sp. (25%), Vismia angustifolia (25%) y Miconia sp. (20%). El resto (Socratea exorrhiza, Apeiba membranacea, Bellucia axinanthera, Castilla elastica, Ficus insipida, Cecropia angustifolia, Ocotea sp. y Brosimum utile) representaron cada una el 5%. Vale la pena resaltar que, de las 19 especies arbóreas presentes en la parcela, solo tres mostraron reclutamiento permanente durante el período 2015-2017. Durante el período evaluado la mortalidad fue de seis individuos, mientras que el reclutamiento estuvo representado por 30. Lo anterior muestra una relación importante entre estos dos factores porque se mantiene más o menos constante la densidad de árboles de la unidad de muestreo. En este sentido, algunos investigadores (Peralta et al. 1987, Ramírez et al. 2002) afirman que las variables estructurales como densidad, área basal y número de especies, fluctúan en torno a un valor medio a lo largo del tiempo, y que esa constancia relativa es mantenida a través de un balance adecuado entre las tasas de mortalidad y reclutamiento de árboles. Estos procesos hacen que el bosque se conserve en equilibrio dinámico en términos de su biomasa en largos períodos (Bormann y Linkens 1979); sin embargo, otros investigadores (Phillips et al. 1994) afirman que debido al cambio climático global y a procesos como la fragmentación de hábitats, las tasas de mortalidad y reclutamiento de árboles han aumentado en las últimas décadas en muchos bosques tropicales, lo cual supone un incremento en el dinamismo de los bosques tropicales, que podría afectar significativamente la composición y diversidad de especies arbóreas presentes en estos ecosistemas (Quinto et al. 2009). De acuerdo con Swaine et al. (1987) el reclutamiento aquí registrado es una manifestación de la fecundidad de las especies y del crecimiento y sobrevivencia de los juveniles, constituyendo en uno de los aspectos dinámicos más importantes de un ecosistema. Aunque en la mayoría de los reportes provenientes de otros bosques tropicales las tasas de reclutamiento de los árboles están relacionada con la mortalidad, manteniendo más o menos constante la densidad de árboles con DAP≥10 cm, la relación entre mortalidad y reclutamiento puede ser débil para períodos cortos y áreas pequeñas, tal y como sucedió en este estudio, donde el intervalo del muestreo fue de un año y el área representada por 0,25 ha. Conclusiones No se evidenciaron cambios relevantes en la estructura y composición de la vegetación arbórea durante el período monitoreado, pues el promedio diamétrico mostró una variación de 1 cm; situación similar se presentó en la composición, donde solo se incorporaron dos especies (Apeiba membranacea y Brosimum utile) con la mínima cantidad de individuos. El componente arbóreo asociado con la parcela de restauración pasiva, muestran una tendencia muy similar a la reportada en otros ecosistemas afectados por la actividad ganadera, donde convergen las familias Moraceae, Cecropiaceae, Clusiaceae, Melastomataceae, Hypericaceae, Malvaceae y Urticaceae y predominan las especies Vismia angustifolia (punta de lanza), Jacaranda sp. (gualanday o pinguasí), Lacistema aggregatum (café de monte), Himatanthus articulatus (mamancallo), Miconia sp. (uvita o morita), Inga sp. (guamo) y Socratea exorrhiza (palma zancona). Los resultados arrojados a lo largo de estos tres muestreos, reflejan que a pesar de la poca variación en los parámetros estructurales y de composición, las familias y especies aumentan conforme avanza el estadío sucesional, demostrando la tendencia de recuperación de atributos de diversidad, composición y funcionalidad de este ecosistema. A pesar de las muertes presentadas en las especies que inicialmente fueron las más abundantes, estas aún continúan siendo dominantes a razón del número de individuos reclutados durante este tiempo. En términos generales, los resultados obtenidos en esta investigación muestran que el bosque secundario se encuentra en proceso de recuperación, pues la mayor parte de los individuos están concentrados en la clase diamétrica de menor tamaño. Agradecimientos A los coinvestigadores Blas Antonio Cárdenas, Lili Johana Perea Rentería, Yadiris Palacios Mosquera y Carlos Arturo Castaño Muñoz por su apoyo y acompañamiento en el levantamiento de la información de campo. Al Consejo Comunitario de Santa Cecilia y la Asociación de Amigos de la Fauna y la Flora de Santa Cecilia (ASOAFA) por facilitarnos y permitirnos desarrollar la investigación en áreas de su título colectivo. A nivel institucional, al Instituto de Investigaciones Ambientales del Pacífico (IIAP) y a la Corporación Autónoma Regional de Risaralda (CARDER) por su apoyo técnico y financiero. Literatura citada
|